How Severe Anaemia Might Influence the Risk of Invasive Bacterial Infections in African Children

 , , and
, , and {kind=link}
{kind=link}
Abstract
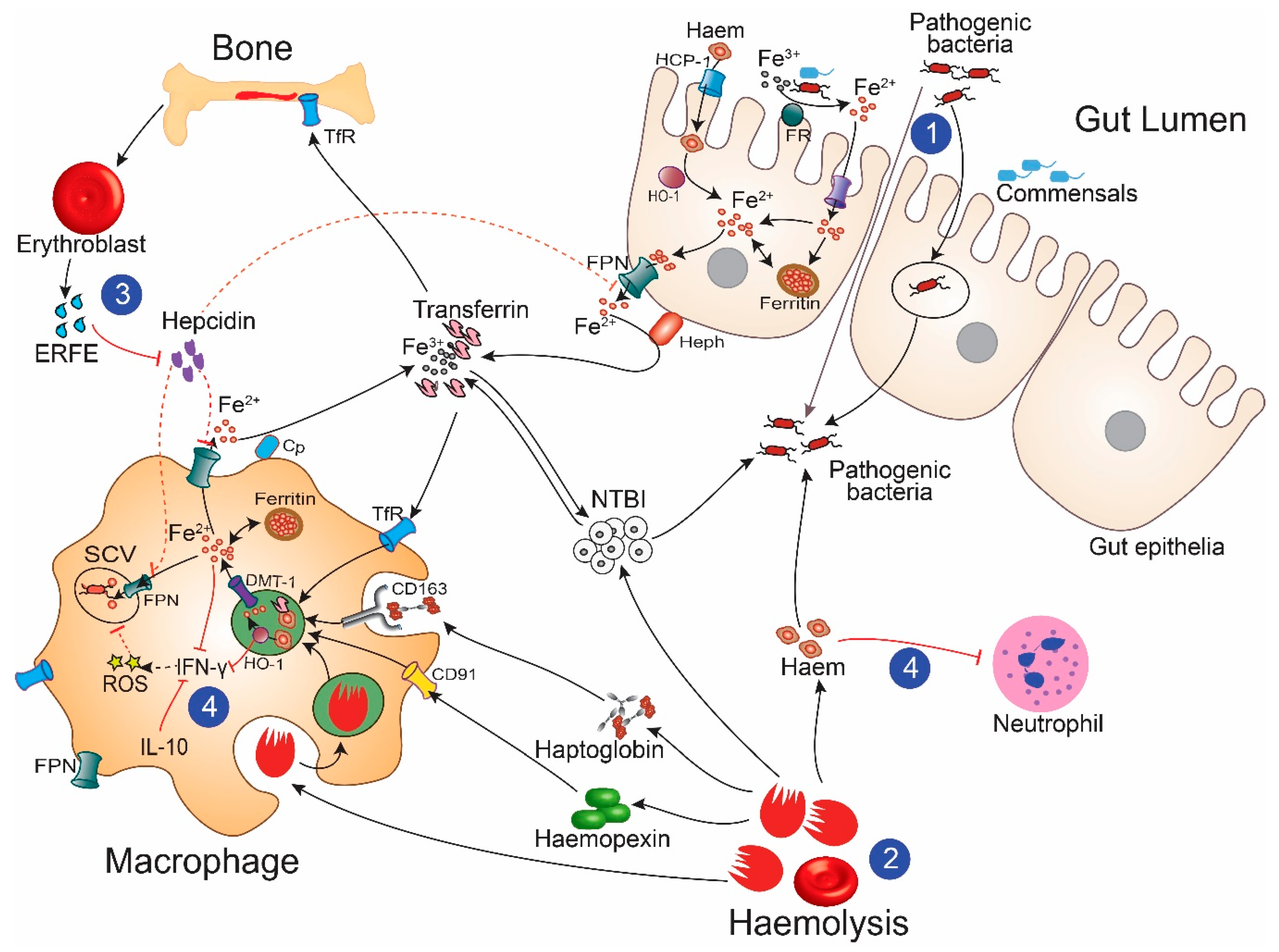
:1. Introduction
2. The Severe Anaemia—Bacteraemia Hypothesis
2.1. Gram-Negative Organisms
2.1.1. Non-Typhoidal Salmonellae
2.1.2. Escherichia coli
2.1.3. Haemophilus influenzae
2.1.4. Other Gram-Negative Organisms
2.2. Gram-Positive Organisms
2.2.1. Staphylococcus aureus
2.2.2. Streptococcus pneumoniae
3. Testing the Hypothesis
3.1. In Vitro and Animal Model Studies
3.2. Observational Studies
3.3. Mendelian Randomization
4. Perspectives and Implications of the Hypothesis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| ERFE | Erythroferrone |
| HO-1 | Haem oxygenase 1 |
| HIV | Human immunodeficiency virus |
| IL | Interleukin |
| IFN-γ | Interferon-gamma |
| MR | Mendelian randomization |
| NTBI | Non-transferrin bound iron |
| NTS | Non-typhoidal Salmonella |
| RCTs | Randomized clinical trials |
| ROS | Reactive oxygen species |
| SCV | Salmonella-containing vacuole |
References
- Liu, L.; Oza, S.; Hogan, D.; Perin, J.; Rudan, I.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of child mortality in 2000–13, with projections to inform post-2015 priorities: An updated systematic analysis. Lancet 2015, 385, 430–440. [Google Scholar] [CrossRef]
- Stevens, G.A.; Finucane, M.M.; De-Regil, L.M.; Paciorek, C.J.; Flaxman, S.R.; Branca, F.; Pena-Rosas, J.P.; Bhutta, Z.A.; Ezzati, M. Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011: A systematic analysis of population-representative data. Lancet Glob. Health 2013, 1, e16–e25. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, R.J.; Clements, A.C. Spatial heterogeneity of haemoglobin concentration in preschool-age children in sub-Saharan Africa. Bull. World Health Organ. 2011, 89, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Kassebaum, N.J. The Global Burden of Anemia. Hematol. Oncol. Clin. N. Am. 2016, 30, 247–308. [Google Scholar] [CrossRef] [Green Version]
- English, M.; Ahmed, M.; Ngando, C.; Berkley, J.; Ross, A. Blood transfusion for severe anaemia in children in a Kenyan hospital. Lancet 2002, 359, 494–495. [Google Scholar] [CrossRef]
- Pedro, R.; Akech, S.; Fegan, G.; Maitland, K. Changing trends in blood transfusion in children and neonates admitted in Kilifi District Hospital, Kenya. Malar. J. 2010, 9, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simbauranga, R.H.; Kamugisha, E.; Hokororo, A.; Kidenya, B.R.; Makani, J. Prevalence and factors associated with severe anaemia amongst under-five children hospitalized at Bugando Medical Centre, Mwanza, Tanzania. BMC Hematol. 2015, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Foote, E.M.; Sullivan, K.M.; Ruth, L.J.; Oremo, J.; Sadumah, I.; Williams, T.N.; Suchdev, P.S. Determinants of anemia among preschool children in rural, western Kenya. Am. J. Trop. Med. Hyg. 2013, 88, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Calis, J.C.; Phiri, K.S.; Faragher, E.B.; Brabin, B.J.; Bates, I.; Cuevas, L.E.; de Haan, R.J.; Phiri, A.I.; Malange, P.; Khoka, M.; et al. Severe anemia in Malawian children. N. Engl. J. Med. 2008, 358, 888–899. [Google Scholar] [CrossRef] [Green Version]
- Reddy, E.A.; Shaw, A.V.; Crump, J.A. Community-acquired bloodstream infections in Africa: A systematic review and meta-analysis. Lancet Infect. Dis. 2010, 10, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Berkley, J.A.; Lowe, B.S.; Mwangi, I.; Williams, T.; Bauni, E.; Mwarumba, S.; Ngetsa, C.; Slack, M.P.; Njenga, S.; Hart, C.A.; et al. Bacteremia among children admitted to a rural hospital in Kenya. N. Engl. J. Med. 2005, 352, 39–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brent, A.J.; Ahmed, I.; Ndiritu, M.; Lewa, P.; Ngetsa, C.; Lowe, B.; Bauni, E.; English, M.; Berkley, J.A.; Scott, J.A. Incidence of clinically significant bacteraemia in children who present to hospital in Kenya: Community-based- observational study. Lancet 2006, 367, 482–488. [Google Scholar] [CrossRef]
- Popoola, O.; Kehinde, A.; Ogunleye, V.; Adewusi, O.J.; Toy, T.; Mogeni, O.D.; Aroyewun, E.O.; Agbi, S.; Adekanmbi, O.; Adepoju, A.; et al. Bacteremia Among Febrile Patients Attending Selected Healthcare Facilities in Ibadan, Nigeria. Clin. Infect. Dis. 2019, 69, S466–S473. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.; Beris, P. Pathophysiology and Differential Diagnosis of Anaemia. In The Handbook on Disorders of Erythropoiesis, Erythrocytes and Iron Metabolism; Beaumont, C., Béris, P., Beuzard, Y., Brugnara, C., Eds.; European School of Haematology: Paris, France, 2009; pp. 108–141. [Google Scholar]
- van Hensbroek, M.B.; Jonker, F.; Bates, I. Severe acquired anaemia in Africa: New concepts. Br. J. Haematol. 2011, 154, 690–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundd, P.; Gladwin, M.T.; Novelli, E.M. Pathophysiology of Sickle Cell Disease. Annu. Rev. Pathol. 2019, 14, 263–292. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Anemia of inflammation. Hematol. Oncol. Clin. N. Am. 2014, 28, 671–681. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, E.; Rivera, S.; Gabayan, V.; Keller, C.; Taudorf, S.; Pedersen, B.K.; Ganz, T. IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. J. Clin. Investig. 2004, 113, 1271–1276. [Google Scholar] [CrossRef] [Green Version]
- Arezes, J.; Jung, G.; Gabayan, V.; Valore, E.; Ruchala, P.; Gulig, P.A.; Ganz, T.; Nemeth, E.; Bulut, Y. Hepcidin-induced hypoferremia is a critical host defense mechanism against the siderophilic bacterium Vibrio vulnificus. Cell Host Microbe 2015, 17, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Z.; He, Y.X.; Yang, C.J.; Zhou, W.; Zou, C.G. Hepcidin is regulated during blood-stage malaria and plays a protective role in malaria infection. J. Immunol. 2011, 187, 6410–6416. [Google Scholar] [CrossRef] [Green Version]
- Lokken, K.L.; Stull-Lane, A.R.; Poels, K.; Tsolis, R.M. Malaria Parasite-Mediated Alteration of Macrophage Function and Increased Iron Availability Predispose to Disseminated Nontyphoidal Salmonella Infection. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Nadjm, B.; Amos, B.; Mtove, G.; Ostermann, J.; Chonya, S.; Wangai, H.; Kimera, J.; Msuya, W.; Mtei, F.; Dekker, D.; et al. WHO guidelines for antimicrobial treatment in children admitted to hospital in an area of intense Plasmodium falciparum transmission: Prospective study. BMJ 2010, 340, c1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigauque, B.; Roca, A.; Mandomando, I.; Morais, L.; Quinto, L.; Sacarlal, J.; Macete, E.; Nhamposa, T.; Machevo, S.; Aide, P.; et al. Community-acquired bacteremia among children admitted to a rural hospital in Mozambique. Pediatr. Infect. Dis. J. 2009, 28, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Brent, A.J.; Oundo, J.O.; Mwangi, I.; Ochola, L.; Lowe, B.; Berkley, J.A. Salmonella bacteremia in Kenyan children. Pediatr. Infect. Dis. J. 2006, 25, 230–236. [Google Scholar] [CrossRef]
- Graham, S.M.; Walsh, A.L.; Molyneux, E.M.; Phiri, A.J.; Molyneux, M.E. Clinical presentation of non-typhoidal Salmonella bacteraemia in Malawian children. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 310–314. [Google Scholar] [CrossRef]
- Biggs, H.M.; Lester, R.; Nadjm, B.; Mtove, G.; Todd, J.E.; Kinabo, G.D.; Philemon, R.; Amos, B.; Morrissey, A.B.; Reyburn, H.; et al. Invasive Salmonella infections in areas of high and low malaria transmission intensity in Tanzania. Clin. Infect. Dis. 2014, 58, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Bronzan, R.N.; Taylor, T.E.; Mwenechanya, J.; Tembo, M.; Kayira, K.; Bwanaisa, L.; Njobvu, A.; Kondowe, W.; Chalira, C.; Walsh, A.L.; et al. Bacteremia in Malawian children with severe malaria: Prevalence, etiology, HIV coinfection, and outcome. J. Infect. Dis. 2007, 195, 895–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, S.M.; Mwenechanya, J.; Tembo, M.; Kabudula, M.; Molyneux, E.M.; Walsh, A.L.; Molyneux, M.E.; Taylor, T.E. The pattern of bacteraemia in children with severe malaria. Malawi Med. J. 2002, 14, 11–15. [Google Scholar]
- Church, J.; Maitland, K. Invasive bacterial co-infection in African children with Plasmodium falciparum malaria: A systematic review. BMC Med. 2014, 12, 31. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.N.; Uyoga, S.; Macharia, A.; Ndila, C.; McAuley, C.F.; Opi, D.H.; Mwarumba, S.; Makani, J.; Komba, A.; Ndiritu, M.N.; et al. Bacteraemia in Kenyan children with sickle-cell anaemia: A retrospective cohort and case-control study. Lancet 2009, 374, 1364–1370. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, M.; Moïsi, J.C.; Klugman, K.P.; Iglesias, J.M.; Grant, L.R.; Mpoudi-Etame, M.; Levine, O.S. Increased risk of invasive bacterial infections in African people with sickle-cell disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2010, 10, 329–337. [Google Scholar] [CrossRef]
- White, N.J. Anaemia and malaria. Malar. J. 2018, 17, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunnington, A.J.; de Souza, J.B.; Walther, M.; Riley, E.M. Malaria impairs resistance to Salmonella through heme- and heme oxygenase-dependent dysfunctional granulocyte mobilization. Nat. Med. 2011, 18, 120–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, C.M.; Butler, B.P.; Chau, J.Y.; Paixao, T.A.; Cheung, K.W.; Santos, R.L.; Luckhart, S.; Tsolis, R.M. Both hemolytic anemia and malaria parasite-specific factors increase susceptibility to Nontyphoidal Salmonella enterica serovar typhimurium infection in mice. Infect. Immunol. 2010, 78, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Kaye, D.; Hook, E.W. The influence of hemolysis on susceptibility to Salmonella infection: Additional observations. J. Immunol. 1963, 91, 518–527. [Google Scholar] [PubMed]
- Kaye, D.; Merselis, J.G., Jr.; Hook, E.W. Influence of Plasmodium berghei infection on susceptibility to salmonella infection. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1965, 120, 810–813. [Google Scholar] [CrossRef]
- Jonker, F.A.; Calis, J.C.; Phiri, K.; Kraaijenhagen, R.J.; Brabin, B.J.; Faragher, B.; Wiegerinck, E.T.; Tjalsma, H.; Swinkels, D.W.; van Hensbroek, M.B. Low hepcidin levels in severely anemic malawian children with high incidence of infectious diseases and bone marrow iron deficiency. PLoS ONE 2013, 8, e78964. [Google Scholar] [CrossRef]
- Kautz, L.; Jung, G.; Valore, E.V.; Rivella, S.; Nemeth, E.; Ganz, T. Identification of erythroferrone as an erythroid regulator of iron metabolism. Nat. Genet. 2014, 46, 678–684. [Google Scholar] [CrossRef] [Green Version]
- Latour, C.; Wlodarczyk, M.F.; Jung, G.; Gineste, A.; Blanchard, N.; Ganz, T.; Roth, M.P.; Coppin, H.; Kautz, L. Erythroferrone contributes to hepcidin repression in a mouse model of malarial anemia. Haematologica 2017, 102, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Makani, J.; Tluway, F.; Makubi, A.; Armitage, A.E.; Pasricha, S.R.; Drakesmith, H.; Prentice, A.M.; Cox, S.E. Decreased Hepcidin Levels Are Associated with Low Steady-state Hemoglobin in Children With Sickle Cell Disease in Tanzania. EBioMedicine 2018, 34, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Weiss, G. Iron in infection and immunity. Mol. Asp. Med. 2020. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron homeostasis in host defence and inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engle-Stone, R.; Aaron, G.J.; Huang, J.; Wirth, J.P.; Namaste, S.M.; Williams, A.M.; Peerson, J.M.; Rohner, F.; Varadhan, R.; Addo, O.Y.; et al. Predictors of anemia in preschool children: Biomarkers Reflecting Inflammation and Nutritional Determinants of Anemia (BRINDA) project. Am. J. Clin. Nutr. 2017, 106, 402s–415s. [Google Scholar] [CrossRef] [PubMed]
- Newton, C.R.; Warn, P.A.; Winstanley, P.A.; Peshu, N.; Snow, R.W.; Pasvol, G.; Marsh, K. Severe anaemia in children living in a malaria endemic area of Kenya. Trop. Med. Int. Health 1997, 2, 165–178. [Google Scholar] [CrossRef]
- Adegoke, S.; Ayansanwo, A.; Oluwayemi, I.; Okeniyi, J. Determinants of mortality in Nigerian children with severe anaemia. S. Afr. Med. J. 2012, 102, 807–810. [Google Scholar] [CrossRef]
- Muoneke, V.; ChidiIbekwe, R. Prevalence and aetiology of severe anaemia in under-5 children in Abakaliki South Eastern Nigeria. Pediatr. Ther. 2011, 1, 107. [Google Scholar] [CrossRef]
- Nyakeriga, A.M.; Troye-Blomberg, M.; Dorfman, J.R.; Alexander, N.D.; Back, R.; Kortok, M.; Chemtai, A.K.; Marsh, K.; Williams, T.N. Iron deficiency and malaria among children living on the coast of Kenya. J. Infect. Dis. 2004, 190, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Muriuki, J.M.; Mentzer, A.J.; Kimita, W.; Ndungu, F.M.; Macharia, A.W.; Webb, E.L.; Lule, S.A.; Morovat, A.; Hill, A.V.S.; Bejon, P.; et al. Iron Status and Associated Malaria Risk Among African Children. Clin. Infect. Dis. 2019, 68, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Michels, K.R.; Lambrecht, N.J.; Carson, W.F.; Schaller, M.A.; Lukacs, N.W.; Bermick, J.R. The Role of Iron in the Susceptibility of Neonatal Mice to Escherichia coli K1 Sepsis. J. Infect. Dis. 2019, 220, 1219–1229. [Google Scholar] [CrossRef]
- Hassan, T.H.; Badr, M.A.; Karam, N.A.; Zkaria, M.; El Saadany, H.F.; Rahman, D.M.; Shahbah, D.A.; Al Morshedy, S.M.; Fathy, M.; Esh, A.M.; et al. Impact of iron deficiency anemia on the function of the immune system in children. Medicine 2016, 95, e5395. [Google Scholar] [CrossRef]
- Darshan, D.; Frazer, D.M.; Wilkins, S.J.; Anderson, G.J. Severe iron deficiency blunts the response of the iron regulatory gene Hamp and pro-inflammatory cytokines to lipopolysaccharide. Haematologica 2010, 95, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Li, C.; Wu, Q.; An, P.; Huang, L.; Wang, J.; Chen, C.; Chen, X.; Zhang, F.; Ma, L.; et al. Iron-dependent histone 3 lysine 9 demethylation controls B cell proliferation and humoral immune responses. Nat. Commun. 2019, 10, 2935. [Google Scholar] [CrossRef] [Green Version]
- Das, I.; Saha, K.; Mukhopadhyay, D.; Roy, S.; Raychaudhuri, G.; Chatterjee, M.; Mitra, P.K. Impact of iron deficiency anemia on cell-mediated and humoral immunity in children: A case control study. J. Nat. Sci. Biol. Med. 2014, 5, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochim. Biophys. Acta 2012, 1823, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Mohus, R.M.; Paulsen, J.; Gustad, L.; Askim, A.; Mehl, A.; DeWan, A.T.; Afset, J.E.; Asvold, B.O.; Solligard, E.; Damas, J.K. Association of iron status with the risk of bloodstream infections: Results from the prospective population-based HUNT Study in Norway. Intensive Care Med. 2018, 44, 1276–1283. [Google Scholar] [CrossRef]
- Swenson, E.R.; Porcher, R.; Piagnerelli, M. Iron deficiency and infection: Another pathway to explore in critically ill patients? Intensive Care Med. 2018, 44, 2260–2262. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Dichtl, S.; Schroll, A.; Haschka, D.; Tymoszuk, P.; Theurl, I.; Weiss, G. Iron and innate antimicrobial immunity-Depriving the pathogen, defending the host. J. Trace Elem. Med. Biol. 2018, 48, 118–133. [Google Scholar] [CrossRef]
- de Bruin, A.M.; Voermans, C.; Nolte, M.A. Impact of interferon-gamma on hematopoiesis. Blood 2014, 124, 2479–2486. [Google Scholar] [CrossRef] [Green Version]
- Mandala, W.L.; Msefula, C.L.; Gondwe, E.N.; Drayson, M.T.; Molyneux, M.E.; MacLennan, C.A. Cytokine Profiles in Malawian Children Presenting with Uncomplicated Malaria, Severe Malarial Anemia, and Cerebral Malaria. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef] [Green Version]
- Burte, F.; Brown, B.J.; Orimadegun, A.E.; Ajetunmobi, W.A.; Afolabi, N.K.; Akinkunmi, F.; Kowobari, O.; Omokhodion, S.; Osinusi, K.; Akinbami, F.O.; et al. Circulatory hepcidin is associated with the anti-inflammatory response but not with iron or anemic status in childhood malaria. Blood 2013, 121, 3016–3022. [Google Scholar] [CrossRef]
- Stoffel, N.U.; Lazrak, M.; Bellitir, S.; Mir, N.E.; Hamdouchi, A.E.; Barkat, A.; Zeder, C.; Moretti, D.; Aguenaou, H.; Zimmermann, M.B. The opposing effects of acute inflammation and iron deficiency anemia on serum hepcidin and iron absorption in young women. Haematologica 2019, 104, 1143–1149. [Google Scholar] [CrossRef] [Green Version]
- Berkley, J.A.; Bejon, P.; Mwangi, T.; Gwer, S.; Maitland, K.; Williams, T.N.; Mohammed, S.; Osier, F.; Kinyanjui, S.; Fegan, G.; et al. HIV infection, malnutrition, and invasive bacterial infection among children with severe malaria. Clin. Infect. Dis. 2009, 49, 336–343. [Google Scholar] [CrossRef]
- Lewis, D.K.; Whitty, C.J.M.; Walsh, A.L.; Epino, H.; van den Broek, N.R.; Letsky, E.A.; Munthali, C.; Mukiibi, J.M.; Boeree, M.J. Treatable factors associated with severe anaemia in adults admitted to medical wards in Blantyre, Malawi, an area of high HIV seroprevalence. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 561–567. [Google Scholar] [CrossRef]
- Page, A.L.; de Rekeneire, N.; Sayadi, S.; Aberrane, S.; Janssens, A.C.; Rieux, C.; Djibo, A.; Manuguerra, J.C.; Ducou-le-Pointe, H.; Grais, R.F.; et al. Infections in children admitted with complicated severe acute malnutrition in Niger. PLoS ONE 2013, 8, e68699. [Google Scholar] [CrossRef] [Green Version]
- Mandomando, I.; Macete, E.; Sigauque, B.; Morais, L.; Quinto, L.; Sacarlal, J.; Espasa, M.; Valles, X.; Bassat, Q.; Aide, P.; et al. Invasive non-typhoidal Salmonella in Mozambican children. Trop. Med. Int. Health 2009, 14, 1467–1474. [Google Scholar] [CrossRef]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688s–1693s. [Google Scholar] [CrossRef] [Green Version]
- Paganini, D.; Jaeggi, T.; Cercamondi, C.; Kujinga, P.; Moretti, D.; Zimmermann, M. Anemia and iron status are predictors of gut microbiome composition and metabolites in infants and children in rural Kenya. FASEB J. Conf. Exp. Biol. 2016, 30, 296.2. [Google Scholar]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.L.; Richardson, S.H.; Failla, M.L. Elevated iron status increases bacterial invasion and survival and alters cytokine/chemokine mRNA expression in Caco-2 human intestinal cells. J. Nutr. 2001, 131, 1452–1458. [Google Scholar] [CrossRef]
- MohanKumar, K.; Namachivayam, K.; Sivakumar, N.; Alves, N.G.; Sidhaye, V.; Das, J.K.; Chung, Y.; Breslin, J.W.; Maheshwari, A. Severe neonatal anemia increases intestinal permeability by disrupting epithelial adherens junctions. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G705–G716. [Google Scholar] [CrossRef]
- Arthur, C.M.; Nalbant, D.; Feldman, H.A.; Saeedi, B.J.; Matthews, J.; Robinson, B.S.; Kamili, N.A.; Bennett, A.; Cress, G.A.; Sola-Visner, M.; et al. Anemia induces gut inflammation and injury in an animal model of preterm infants. Transfusion 2019, 59, 1233–1245. [Google Scholar] [CrossRef]
- Spees, A.M.; Kingsbury, D.D.; Wangdi, T.; Xavier, M.N.; Tsolis, R.M.; Bäumler, A.J. Neutrophils are a source of gamma interferon during acute Salmonella enterica serovar Typhimurium colitis. Infect. Immun. 2014, 82, 1692–1697. [Google Scholar] [CrossRef] [Green Version]
- Winter, S.E.; Thiennimitr, P.; Winter, M.G.; Butler, B.P.; Huseby, D.L.; Crawford, R.W.; Russell, J.M.; Bevins, C.L.; Adams, L.G.; Tsolis, R.M.; et al. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010, 467, 426–429. [Google Scholar] [CrossRef]
- Liu, J.Z.; Jellbauer, S.; Poe, A.J.; Ton, V.; Pesciaroli, M.; Kehl-Fie, T.E.; Restrepo, N.A.; Hosking, M.P.; Edwards, R.A.; Battistoni, A.; et al. Zinc sequestration by the neutrophil protein calprotectin enhances Salmonella growth in the inflamed gut. Cell Host Microbe 2012, 11, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Goethel, A.; Croitoru, K.; Philpott, D.J. The interplay between microbes and the immune response in inflammatory bowel disease. J. Physiol. 2018, 596, 3869–3882. [Google Scholar] [CrossRef] [Green Version]
- Kruger, P.; Saffarzadeh, M.; Weber, A.N.; Rieber, N.; Radsak, M.; von Bernuth, H.; Benarafa, C.; Roos, D.; Skokowa, J.; Hartl, D. Neutrophils: Between host defence, immune modulation, and tissue injury. PLoS Pathog. 2015, 11, e1004651. [Google Scholar] [CrossRef] [Green Version]
- Gogoi, M.; Shreenivas, M.M.; Chakravortty, D. Hoodwinking the Big-Eater to Prosper: The Salmonella-Macrophage Paradigm. J. Innate Immun. 2019, 11, 289–299. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Westerman, D.A.; Evans, D.; Metz, J. Neutrophil hypersegmentation in iron deficiency anaemia: A case-control study. Br. J. Haematol. 1999, 107, 512–515. [Google Scholar] [CrossRef]
- Berrak, S.G.; Angaji, M.; Turkkan, E.; Canpolat, C.; Timur, C.; Eksioglu-Demiralp, E. The effects of iron deficiency on neutrophil/monocyte apoptosis in children. Cell Prolif. 2007, 40, 741–754. [Google Scholar] [CrossRef]
- Murakawa, H.; Bland, C.E.; Willis, W.T.; Dallman, P.R. Iron deficiency and neutrophil function: Different rates of correction of the depressions in oxidative burst and myeloperoxidase activity after iron treatment. Blood 1987, 69, 1464–1468. [Google Scholar] [CrossRef] [Green Version]
- Abdelmahmuod, E.; Yassin, M.A.; Ahmed, M. Iron Deficiency Anemia-Induced Neutropenia in Adult Female. Cureus 2020, 12, e8899. [Google Scholar] [CrossRef]
- Abuirmeileh, A.; Bahnassi, A.; Abuirmeileh, A. Unexplained chronic leukopenia treated with oral iron supplements. Int. J. Clin. Pharm. 2014, 36, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Mooney, J.P.; Butler, B.P.; Lokken, K.L.; Xavier, M.N.; Chau, J.Y.; Schaltenberg, N.; Dandekar, S.; George, M.D.; Santos, R.L.; Luckhart, S.; et al. The mucosal inflammatory response to non-typhoidal Salmonella in the intestine is blunted by IL-10 during concurrent malaria parasite infection. Mucosal Immunol. 2014, 7, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Mooney, J.P.; Galloway, L.J.; Riley, E.M. Malaria, anemia, and invasive bacterial disease: A neutrophil problem? J. Leukoc. Biol. 2019, 105, 645–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokken, K.L.; Mooney, J.P.; Butler, B.P.; Xavier, M.N.; Chau, J.Y.; Schaltenberg, N.; Begum, R.H.; Muller, W.; Luckhart, S.; Tsolis, R.M. Malaria parasite infection compromises control of concurrent systemic non-typhoidal Salmonella infection via IL-10-mediated alteration of myeloid cell function. PLoS Pathog. 2014, 10, e1004049. [Google Scholar] [CrossRef]
- Pagani, A.; Nai, A.; Corna, G.; Bosurgi, L.; Rovere-Querini, P.; Camaschella, C.; Silvestri, L. Low hepcidin accounts for the proinflammatory status associated with iron deficiency. Blood 2011, 118, 736–746. [Google Scholar] [CrossRef]
- Kaplanski, G.; Marin, V.; Montero-Julian, F.; Mantovani, A.; Farnarier, C. IL-6: A regulator of the transition from neutrophil to monocyte recruitment during inflammation. Trends Immunol. 2003, 24, 25–29. [Google Scholar] [CrossRef]
- Fielding, C.A.; McLoughlin, R.M.; McLeod, L.; Colmont, C.S.; Najdovska, M.; Grail, D.; Ernst, M.; Jones, S.A.; Topley, N.; Jenkins, B.J. IL-6 regulates neutrophil trafficking during acute inflammation via STAT3. J. Immunol. 2008, 181, 2189–2195. [Google Scholar] [CrossRef]
- Martins, R.; Maier, J.; Gorki, A.D.; Huber, K.V.; Sharif, O.; Starkl, P.; Saluzzo, S.; Quattrone, F.; Gawish, R.; Lakovits, K.; et al. Heme drives hemolysis-induced susceptibility to infection via disruption of phagocyte functions. Nat. Immunol. 2016, 17, 1361–1372. [Google Scholar] [CrossRef]
- Mitterstiller, A.M.; Haschka, D.; Dichtl, S.; Nairz, M.; Demetz, E.; Talasz, H.; Soares, M.P.; Einwallner, E.; Esterbauer, H.; Fang, F.C.; et al. Heme oxygenase 1 controls early innate immune response of macrophages to Salmonella Typhimurium infection. Cell. Microbiol. 2016, 18, 1374–1389. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Basu, M.; Guin, S.; Gupta, P.; Mitterstiller, A.M.; Weiss, G.; Jana, K.; Ukil, A. Leishmania donovani Exploits Macrophage Heme Oxygenase-1 To Neutralize Oxidative Burst and TLR Signaling-Dependent Host Defense. J. Immunol. 2019, 202, 827–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Cui, T.; Chang, Y.; Zhang, W.; Li, S.; He, Y.; Li, B.; Liu, L.; Wang, G.; Gao, T.; et al. HO-1 regulates the function of Treg: Association with the immune intolerance in vitiligo. J. Cell. Mol. Med. 2018, 22, 4335–4343. [Google Scholar] [CrossRef]
- Soares, M.P.; Bach, F.H. Heme oxygenase-1: From biology to therapeutic potential. Trends Mol. Med. 2009, 15, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Werner, E.R.; Werner-Felmayer, G.; Wachter, H. Iron modulates interferon-gamma effects in the human myelomonocytic cell line THP-1. Exp. Hematol. 1992, 20, 605–610. [Google Scholar]
- Oexle, H.; Kaser, A.; Möst, J.; Bellmann-Weiler, R.; Werner, E.R.; Werner-Felmayer, G.; Weiss, G. Pathways for the regulation of interferon-gamma-inducible genes by iron in human monocytic cells. J. Leukoc. Biol. 2003, 74, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Nairz, M.; Fritsche, G.; Brunner, P.; Talasz, H.; Hantke, K.; Weiss, G. Interferon-gamma limits the availability of iron for intramacrophage Salmonella typhimurium. Eur. J. Immunol. 2008, 38, 1923–1936. [Google Scholar] [CrossRef] [PubMed]
- Nairz, M.; Theurl, I.; Ludwiczek, S.; Theurl, M.; Mair, S.M.; Fritsche, G.; Weiss, G. The co-ordinated regulation of iron homeostasis in murine macrophages limits the availability of iron for intracellular Salmonella typhimurium. Cell. Microbiol. 2007, 9, 2126–2140. [Google Scholar] [CrossRef]
- Byrd, T.F. Tumor necrosis factor alpha (TNFalpha) promotes growth of virulent Mycobacterium tuberculosis in human monocytes iron-mediated growth suppression is correlated with decreased release of TNFalpha from iron-treated infected monocytes. J. Clin. Investig. 1997, 99, 2518–2529. [Google Scholar] [CrossRef]
- Melillo, G.; Taylor, L.S.; Brooks, A.; Musso, T.; Cox, G.W.; Varesio, L. Functional requirement of the hypoxia-responsive element in the activation of the inducible nitric oxide synthase promoter by the iron chelator desferrioxamine. J. Biol. Chem. 1997, 272, 12236–12243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noinaj, N.; Guillier, M.; Barnard, T.J.; Buchanan, S.K. TonB-dependent transporters: Regulation, structure, and function. Annu. Rev. Microbiol. 2010, 64, 43–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrow, N.L.; Fleming, R.E.; Minnick, M.F. Sequestration and scavenging of iron in infection. Infect. Immun. 2013, 81, 3503–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischbach, M.A.; Lin, H.; Zhou, L.; Yu, Y.; Abergel, R.J.; Liu, D.R.; Raymond, K.N.; Wanner, B.L.; Strong, R.K.; Walsh, C.T.; et al. The pathogen-associated iroA gene cluster mediates bacterial evasion of lipocalin 2. Proc. Natl. Acad. Sci. USA 2006, 103, 16502–16507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Tamilselvam, B.; Hansen, E.J.; Daefler, S. Modulation of iron homeostasis in macrophages by bacterial intracellular pathogens. BMC Microbiol. 2010, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Boyer, E.; Bergevin, I.; Malo, D.; Gros, P.; Cellier, M.F. Acquisition of Mn(II) in addition to Fe(II) is required for full virulence of Salmonella enterica serovar Typhimurium. Infect. Immun. 2002, 70, 6032–6042. [Google Scholar] [CrossRef] [Green Version]
- Nix, R.N.; Altschuler, S.E.; Henson, P.M.; Detweiler, C.S. Hemophagocytic macrophages harbor Salmonella enterica during persistent infection. PLoS Pathog. 2007, 3, e193. [Google Scholar] [CrossRef]
- Nagy, T.A.; Moreland, S.M.; Detweiler, C.S. Salmonella acquires ferrous iron from haemophagocytic macrophages. Mol. Microbiol. 2014, 93, 1314–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.K.; Jeong, J.H.; Lee, J.M.; Kim, K.S.; Park, S.H.; Kim, Y.D.; Koh, M.; Shin, M.; Jung, Y.S.; Kim, H.S.; et al. Inverse agonist of estrogen-related receptor gamma controls Salmonella typhimurium infection by modulating host iron homeostasis. Nat. Med. 2014, 20, 419–424. [Google Scholar] [CrossRef]
- Nairz, M.; Schleicher, U.; Schroll, A.; Sonnweber, T.; Theurl, I.; Ludwiczek, S.; Talasz, H.; Brandacher, G.; Moser, P.L.; Muckenthaler, M.U.; et al. Nitric oxide-mediated regulation of ferroportin-1 controls macrophage iron homeostasis and immune function in Salmonella infection. J. Exp. Med. 2013, 210, 855–873. [Google Scholar] [CrossRef]
- Chlosta, S.; Fishman, D.S.; Harrington, L.; Johnson, E.E.; Knutson, M.D.; Wessling-Resnick, M.; Cherayil, B.J. The iron efflux protein ferroportin regulates the intracellular growth of Salmonella enterica. Infect. Immunol. 2006, 74, 3065–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.; Kim, K.S.; Jeong, J.H.; Marques, O.; Kim, H.J.; Song, M.; Lee, T.H.; Kim, J.I.; Choi, H.S.; Min, J.J.; et al. The hepcidin-ferroportin axis controls the iron content of Salmonella-containing vacuoles in macrophages. Nat. Commun. 2018, 9, 2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Gan, Z.S.; Ma, W.; Xiong, H.T.; Li, Y.Q.; Wang, Y.Z.; Du, H.H. Synthetic Porcine Hepcidin Exhibits Different Roles in Escherichia coli and Salmonella Infections. Antimicrob. Agents Chemother. 2017, 61, e02638-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuki, K.E.; Eva, M.M.; Richer, E.; Chung, D.; Paquet, M.; Cellier, M.; Canonne-Hergaux, F.; Vaulont, S.; Vidal, S.M.; Malo, D. Suppression of hepcidin expression and iron overload mediate Salmonella susceptibility in ankyrin 1 ENU-induced mutant. PLoS ONE 2013, 8, e55331. [Google Scholar] [CrossRef] [Green Version]
- Casals-Pascual, C.; Huang, H.; Lakhal-Littleton, S.; Thezenas, M.L.; Kai, O.; Newton, C.R.; Roberts, D.J. Hepcidin demonstrates a biphasic association with anemia in acute Plasmodium falciparum malaria. Haematologica 2012, 97, 1695–1698. [Google Scholar] [CrossRef] [Green Version]
- Garénaux, A.; Caza, M.; Dozois, C.M. The Ins and Outs of siderophore mediated iron uptake by extra-intestinal pathogenic Escherichia coli. Vet. Microbiol. 2011, 153, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Otto, B.R.; Sijbrandi, R.; Luirink, J.; Oudega, B.; Heddle, J.G.; Mizutani, K.; Park, S.Y.; Tame, J.R. Crystal structure of hemoglobin protease, a heme binding autotransporter protein from pathogenic Escherichia coli. J. Biol. Chem. 2005, 280, 17339–17345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanova, D.; Raychev, A.; Deville, J.; Humphries, R.; Campeau, S.; Ruchala, P.; Nemeth, E.; Ganz, T.; Bulut, Y. Hepcidin Protects against Lethal Escherichia coli Sepsis in Mice Inoculated with Isolates from Septic Patients. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Cowart, R.E. Reduction of iron by extracellular iron reductases: Implications for microbial iron acquisition. Arch. Biochem. Biophys. 2002, 400, 273–281. [Google Scholar] [CrossRef]
- Cross, J.H.; Bradbury, R.S.; Fulford, A.J.; Jallow, A.T.; Wegmuller, R.; Prentice, A.M.; Cerami, C. Oral iron acutely elevates bacterial growth in human serum. Sci. Rep. 2015, 5, 16670. [Google Scholar] [CrossRef] [Green Version]
- Prentice, S.; Jallow, A.T.; Sinjanka, E.; Jallow, M.W.; Sise, E.A.; Kessler, N.J.; Wegmuller, R.; Cerami, C.; Prentice, A.M. Hepcidin mediates hypoferremia and reduces the growth potential of bacteria in the immediate post-natal period in human neonates. Sci. Rep. 2019, 9, 16596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J. Biol. Chem. 2001, 276, 7806–7810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, D.M.; Reeve, A.W. Increased incidence of gram-negative neonatal sepsis with intramuscula iron administration. Pediatrics 1977, 60, 908–912. [Google Scholar] [PubMed]
- Valdez, J.M.; Scheinberg, P.; Young, N.S.; Walsh, T.J. Infections in patients with aplastic anemia. Semin. Hematol. 2009, 46, 269–276. [Google Scholar] [CrossRef]
- Christopher, G.W. Escherichia coli bacteremia, meningitis, and hemochromatosis. Arch. Intern. Med. 1985, 145, 1908. [Google Scholar] [CrossRef]
- Camaschella, C. Iron and hepcidin: A story of recycling and balance. Hematol. Am. Soc. Hematol. Educ. Program 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fillebeen, C.; Wilkinson, N.; Charlebois, E.; Katsarou, A.; Wagner, J.; Pantopoulos, K. Hepcidin-mediated hypoferremic response to acute inflammation requires a threshold of Bmp6/Hjv/Smad signaling. Blood 2018, 132, 1829–1841. [Google Scholar] [CrossRef] [Green Version]
- Stull, T.L. Protein sources of heme for Haemophilus influenzae. Infect. Immun. 1987, 55, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Morton, D.J.; Seale, T.W.; Bakaletz, L.O.; Jurcisek, J.A.; Smith, A.; VanWagoner, T.M.; Whitby, P.W.; Stull, T.L. The heme-binding protein (HbpA) of Haemophilus influenzae as a virulence determinant. Int. J. Med. Microbiol. 2009, 299, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Morton, D.J.; Whitby, P.W.; Jin, H.; Ren, Z.; Stull, T.L. Effect of multiple mutations in the hemoglobin- and hemoglobin-haptoglobin-binding proteins, HgpA, HgpB, and HgpC, of Haemophilus influenzae type b. Infect. Immun. 1999, 67, 2729–2739. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Arce, I.; Al-Jubair, T.; Euba, B.; Fernández-Calvet, A.; Gil-Campillo, C.; Martí, S.; Törnroth-Horsefield, S.; Riesbeck, K.; Garmendia, J. Moonlighting of Haemophilus influenzae heme acquisition systems contributes to the host airway-pathogen interplay in a coordinated manner. Virulence 2019, 10, 315–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cope, L.D.; Yogev, R.; Muller-Eberhard, U.; Hansen, E.J. A gene cluster involved in the utilization of both free heme and heme:hemopexin by Haemophilus influenzae type b. J. Bacteriol. 1995, 177, 2644–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.G.; Shouldice, S.R.; Kirby, S.D.; Yu, R.H.; Tari, L.W.; Schryvers, A.B. High-affinity binding by the periplasmic iron-binding protein from Haemophilus influenzae is required for acquiring iron from transferrin. Biochem. J. 2007, 404, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shurin, S.B.; Anderson, P.; Zollinger, J.; Rathbun, R.K. Pathophysiology of hemolysis in infections with Hemophilus influenzae type b. J. Clin. Investig. 1986, 77, 1340–1348. [Google Scholar] [CrossRef]
- Kaplan, K.M.; Oski, F.A. Anemia with Haemophilus influenzae meningitis. Pediatrics 1980, 65, 1101–1104. [Google Scholar]
- Kato, G.J.; Steinberg, M.H.; Gladwin, M.T. Intravascular hemolysis and the pathophysiology of sickle cell disease. J. Clin. Investig. 2017, 127, 750–760. [Google Scholar] [CrossRef]
- Saif, M.W. HIV-associated autoimmune hemolytic anemia: An update. AIDS Patient Care STDS 2001, 15, 217–224. [Google Scholar] [CrossRef]
- Stefanova, D.; Raychev, A.; Arezes, J.; Ruchala, P.; Gabayan, V.; Skurnik, M.; Dillon, B.J.; Horwitz, M.A.; Ganz, T.; Bulut, Y.; et al. Endogenous hepcidin and its agonist mediate resistance to selected infections by clearing non-transferrin-bound iron. Blood 2017, 130, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Michels, K.R.; Zhang, Z.; Bettina, A.M.; Cagnina, R.E.; Stefanova, D.; Burdick, M.D.; Vaulont, S.; Nemeth, E.; Ganz, T.; Mehrad, B. Hepcidin-mediated iron sequestration protects against bacterial dissemination during pneumonia. JCI Insight 2017, 2, e92002. [Google Scholar] [CrossRef]
- Lau, C.K.; Krewulak, K.D.; Vogel, H.J. Bacterial ferrous iron transport: The Feo system. FEMS Microbiol. Rev. 2016, 40, 273–298. [Google Scholar] [CrossRef]
- Bracken, C.S.; Baer, M.T.; Abdur-Rashid, A.; Helms, W.; Stojiljkovic, I. Use of heme-protein complexes by the Yersinia enterocolitica HemR receptor: Histidine residues are essential for receptor function. J. Bacteriol. 1999, 181, 6063–6072. [Google Scholar] [CrossRef] [Green Version]
- Noinaj, N.; Easley, N.C.; Oke, M.; Mizuno, N.; Gumbart, J.; Boura, E.; Steere, A.N.; Zak, O.; Aisen, P.; Tajkhorshid, E.; et al. Structural basis for iron piracy by pathogenic Neisseria. Nature 2012, 483, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins-Balding, D.; Baer, M.T.; Stojiljkovic, I. Identification of functionally important regions of a haemoglobin receptor from Neisseria meningitidis. Microbiology 2003, 149, 3423–3435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresso, A.W.; Garufi, G.; Schneewind, O. Bacillus anthracis secretes proteins that mediate heme acquisition from hemoglobin. PLoS Pathog. 2008, 4, e1000132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daou, N.; Buisson, C.; Gohar, M.; Vidic, J.; Bierne, H.; Kallassy, M.; Lereclus, D.; Nielsen-LeRoux, C. IlsA, a unique surface protein of Bacillus cereus required for iron acquisition from heme, hemoglobin and ferritin. PLoS Pathog. 2009, 5, e1000675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, V.J.; Pishchany, G.; Humayun, M.; Schneewind, O.; Skaar, E.P. Staphylococcus aureus IsdB is a hemoglobin receptor required for heme iron utilization. J. Bacteriol. 2006, 188, 8421–8429. [Google Scholar] [CrossRef] [Green Version]
- Skaar, E.P.; Humayun, M.; Bae, T.; DeBord, K.L.; Schneewind, O. Iron-source preference of Staphylococcus aureus infections. Science 2004, 305, 1626–1628. [Google Scholar] [CrossRef] [Green Version]
- Torres, V.J.; Stauff, D.L.; Pishchany, G.; Bezbradica, J.S.; Gordy, L.E.; Iturregui, J.; Anderson, K.L.; Dunman, P.M.; Joyce, S.; Skaar, E.P. A Staphylococcus aureus regulatory system that responds to host heme and modulates virulence. Cell Host Microbe 2007, 1, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Modun, B.; Kendall, D.; Williams, P. Staphylococci express a receptor for human transferrin: Identification of a 42-kilodalton cell wall transferrin-binding protein. Infect. Immun. 1994, 62, 3850–3858. [Google Scholar] [CrossRef] [Green Version]
- Cotton, J.L.; Tao, J.; Balibar, C.J. Identification and characterization of the Staphylococcus aureus gene cluster coding for staphyloferrin A. Biochemistry 2009, 48, 1025–1035. [Google Scholar] [CrossRef]
- Beasley, F.C.; Marolda, C.L.; Cheung, J.; Buac, S.; Heinrichs, D.E. Staphylococcus aureus transporters Hts, Sir, and Sst capture iron liberated from human transferrin by Staphyloferrin A, Staphyloferrin B, and catecholamine stress hormones, respectively, and contribute to virulence. Infect. Immun. 2011, 79, 2345–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, S.S.; Lee, C.J.; Winter, R.E. Hemin utilization is related to virulence of Streptococcus pneumoniae. Infect. Immun. 1993, 61, 5401–5405. [Google Scholar] [CrossRef] [Green Version]
- Romero-Espejel, M.E.; González-López, M.A.; de Jesús Olivares-Trejo, J. Streptococcus pneumoniae requires iron for its viability and expresses two membrane proteins that bind haemoglobin and haem. Metallomics 2013, 5, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.S.; Gilliland, S.M.; Holden, D.W. A Streptococcus pneumoniae pathogenicity island encoding an ABC transporter involved in iron uptake and virulence. Mol. Microbiol. 2001, 40, 572–585. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.S.; Gilliland, S.M.; Ruiz-Albert, J.; Holden, D.W. Characterization of pit, a Streptococcus pneumoniae iron uptake ABC transporter. Infect. Immun. 2002, 70, 4389–4398. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.; Jung, C.L.; Gabayan, V.; Deng, J.C.; Ganz, T.; Nemeth, E.; Bulut, Y. Hepcidin induction by pathogens and pathogen-derived molecules is strongly dependent on interleukin-6. Infect. Immun. 2014, 82, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Morton, D.J.; VanWagoner, T.M.; Seale, T.W.; Whitby, P.W.; Stull, T.L. Differential utilization by Haemophilus influenzae of haemoglobin complexed to the three human haptoglobin phenotypes. FEMS Immunol. Med. Microbiol. 2006, 46, 426–432. [Google Scholar] [CrossRef]
- Pishchany, G.; McCoy, A.L.; Torres, V.J.; Krause, J.C.; Crowe, J.E., Jr.; Fabry, M.E.; Skaar, E.P. Specificity for human hemoglobin enhances Staphylococcus aureus infection. Cell Host Microbe 2010, 8, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Bassat, Q.; Guinovart, C.; Sigauque, B.; Mandomando, I.; Aide, P.; Sacarlal, J.; Nhampossa, T.; Bardaji, A.; Morais, L.; Machevo, S.; et al. Severe malaria and concomitant bacteraemia in children admitted to a rural Mozambican hospital. Trop. Med. Int. Health 2009, 14, 1011–1019. [Google Scholar] [CrossRef]
- Smith, G.D.; Ebrahim, S. ‘Mendelian randomization’: Can genetic epidemiology contribute to understanding environmental determinants of disease? Int. J. Epidemiol. 2003, 32, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, N.A.; Didelez, V.; Burton, P.R.; Tobin, M.D. Mendelian randomisation and causal inference in observational epidemiology. PLoS Med. 2008, 5, e177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Daily Iron Supplementation in Infants and Children. Available online: https://www.who.int/nutrition/publications/micronutrients/guidelines/daily_iron_supp_childrens/en/ (accessed on 4 September 2020).


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abuga, K.M.; Muriuki, J.M.; Williams, T.N.; Atkinson, S.H. How Severe Anaemia Might Influence the Risk of Invasive Bacterial Infections in African Children. Int. J. Mol. Sci. 2020, 21, 6976. https://doi.org/10.3390/ijms21186976
Abuga KM, Muriuki JM, Williams TN, Atkinson SH. How Severe Anaemia Might Influence the Risk of Invasive Bacterial Infections in African Children. International Journal of Molecular Sciences. 2020; 21(18):6976. https://doi.org/10.3390/ijms21186976
Chicago/Turabian StyleAbuga, Kelvin M., John Muthii Muriuki, Thomas N. Williams, and Sarah H. Atkinson. 2020. "How Severe Anaemia Might Influence the Risk of Invasive Bacterial Infections in African Children" International Journal of Molecular Sciences 21, no. 18: 6976. https://doi.org/10.3390/ijms21186976